- Home
- Research Interests Apri sottomenù
- Publications
- People
-
Projects
Apri sottomenù
- HORIZON EUROPE MSCA DN - EXPLORA: Perception of materials, objects and spaces through active EXPLORAtion
- Progetto PNC - Fit4MedRob - Interfacce Neurorobotiche Ibride per il Controllo e il Recupero della Funzionalità Motoria e Verbale (NeuroRobCoRe)
- Progetto PNC - Fit4MedRob - Sistema innovativo per l'Analisi del Movimento e il Monitoraggio in Tempo Reale del Tremore (SMART-tremor)
- PRIN2022 - MulWalk
- Partenariato Esteso, funded by PNRR - Mnesys
- MSCA Staff Exchanges - PLACES
- PRIN 2020
- FETPROACT-2019 MAIA
- HumaneAINet
- Ricerca Finalizzata 2019 - GR-2019-12369242
- PRIN2017 - PACE
- H2020-MSCA-RISE-2016 - Platypus
- Agenda
Human neurophysiology of goal-directed behaviors
Imaging studies
(in collaboration with Prof.ssa, S. Pitzalis, University of Rome, Italy, Prof. M.Sereno, UCSF, USA, and J. Culham, University of Western Ontario, Canada).
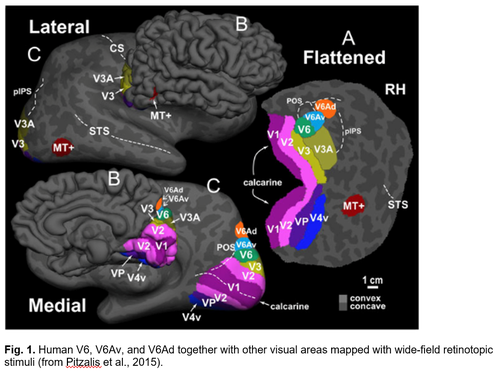
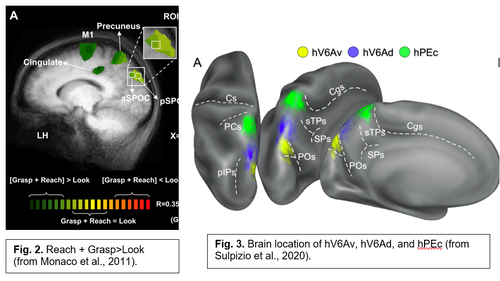
In a series of fMRI investigations we were able to find the human homologues of macaque areas V6, V6Av, and V6Ad, in the medial parieto-occipital cortex (Fig. 1). Similar to the macaque counterparts, the human V6 (hV6) and V6Av (hV6Av), that we called together hV6+, are visual motion areas representing the entire visual field up to the very far periphery. They are very sensitive to the direction of movement of objects in the visual field and to an optical flow similar to that evoked by self-motion. The hV6Ad, instead, again like to the macaque counterpart, is a visuomotor area involved in the control of arm movements in reach-to-grasp actions (Fig. 2). More recently, in the caudal part of the superior parietal lobule, anterior to hV6Ad, we have also found the homologue of macaque areas PEc (Fig. 3). Like in the macaque, the hPEc is activated by movements of both upper and lower limbs.
TMS experiments
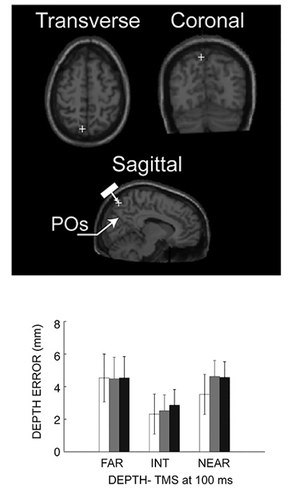
In collaboration with our colleagues Prof. Avenanti, Borgomaneri, Tessari of the Department of Psychology here at UNIBO, we are investigating the causal role of area V6A in humans with TMS during reaching movements. We have found that TMS delivered over hV6A during reach planning affected the encoding of the depth of reaching target by decreasing the accuracy of movements towards targets located farther with respect to the gazed position, but only when they were also far from the body1 (fig. 1). The effectiveness of both retinotopic (farther with respect to the gaze) and spatial position (far from the body) is in agreement with the presence in the monkey V6A of neurons employing either retinotopic, spatial, or mixed reference frames during reach plan. This work provides the first causal evidence of the critical role of hV6A in the planning of visually-guided reaching movements in depth, and represents an example of the transfer of knowledge acquired from studies in the monkey to the human brain.
Breveglieri R., Bosco A., Borgomaneri S., Tessari A., Galletti C., Avenanti A., Fattori P. Transcranial magnetic stimulation over the human medial posterior parietal cortex disrupts depth encoding during reach planning. Cerebral Cortex, February 2020;00:1-14. doi:10.1093/cercor/bhaa224.
References Imagining studies
Pitzalis S., Galletti C., Huang R. S., Patria F., Committeri G., Galati G., Fattori P., and Sereno M. I. (2006) Wide-Field Retinotopy Defines Human Cortical Visual Area V6 J. Neurosci., 26:7962-7973.
Pitzalis S., Sereno M. I., Committeri G., Fattori P., Galati G., Patria F., Galletti C., (2010) Human V6: The Medial Motion Area. Cereb. Cortex, 20:411-424.
Cavina-Pratesi C., Monaco S., Fattori P., Galletti C., McAdam T.D., Quinlan D.J., Goodale M.A., Culham J.C. (2010) Functional magnetic resonance imaging reveals the neural substrates of arm transport and grip formation in reach-to-grasp actions in humans. J Neurosci., 30:10306-23.
Monaco S., Cavina-Pratesi C., Sedda A., Fattori P., Galletti C., Culham JC., (2011). Functional magnetic resonance adaptation reveals the involvement of the dorsomedial stream in hand orientation for grasping. J Neurophysiol., 106:2248-63.
Pitzalis S., Bozzacchi C., Bultrini A., Fattori P., Galletti C., Di Russo F., (2013). Parallel motion signals to the medial and lateral motion areas V6 and MT+. Neuroimage, 67: 89–100.
Pitzalis S, Fattori P., Galletti C., (2013). The functional role of the medial motion area V6. Front. Behav. Neurosci., 6, doi: 10.3389/fnbeh.2012.00091.
Pitzalis S., Sdoia S., Bultrini A., Committeri G., Di Russo F., Fattori P., Galletti C., Galati G., (2013). Selectivity to translational egomotion in human brain motion areas. PLoS One., 8:e60241.
Pitzalis S., Sereno M.I., Committeri G., Fattori P., Galati G., Tosoni A., Galletti C., (2013). The human homologue of macaque area V6A. NeuroImage, 82:517–530.
Tosoni A., Pitzalis S., Committeri G., Fattori P., Galletti C., Galati G., (2015). Resting-state connectivity and functional specialization in human medial parieto-occipital cortex. Brain Struct Funct, 220:3307-21.
Pitzalis S., Fattori P., Galati G., Galletti C., (2015). The human cortical areas V6 and V6A.Visual Neuroscience, 32:e007.
Pitzalis S., Serra C., Sulpizio V., Di Marco S:, Fattori P:, Galati G., Galletti C., (2019). A putative human homologue of the macaque area PEc. NeuroImage, 202:116092.
Serra C., Galletti C., Di Marco S., Fattori P., Galati G., Sulpizio V., Pitzalis S., (2019). Egomotion-related visual areas respond to active leg movements. Hum Brain Mapp., 40:3174–3191.
Sulpizio V., N eri A., Fattori P., Galletti C., Pitzalis S., Galati G., (2020). Real and Imagined Grasping Movements Differently Activate the Human Dorsomedial Parietal Cortex. Neuroscience, 434:22–34.
Sulpizio V., Galati G., Fattori P., Galletti C., Pitzalis S., (2020). A common neural substrate for processing scenes and egomotion‐compatible visual motion. Brain Structure and Function, 225:2091–2110.
Pitzalis S., Serra C:, Sulpizio V., Committeri G:, de Pasquale F., Fattori P., Galletti C., Sepe R., Galati G., (2020). Neural bases of self- and object-motion in a naturalistic vision. Hum Brain Mapp., 41:1084–1111.